Zygomycota
| zygomycota | |
|---|---|
 | |
| Phycomyces sp。 | |
| 科學分類 | |
| 領域: | 真核生物 |
| 王國: | 菌類 |
| 分配: | zygomycota C. Moreau 1954(非正式) |
| 班級: | Zygomycetes 1881年冬 |
| 命令 | |
粘膜菌: 昆蟲術: | |
Zygomycota或Zygote Fungi是王國真菌的前部或門。現在,成員是兩個門的一部分:粘膜瘤和浮游動物。已知大約1060種。它們大多在棲息地,生活在土壤或腐爛的植物或動物材料中。有些是植物,昆蟲和小動物的寄生蟲,而另一些則與植物形成共生關係。合黴菌菌絲可以是凝聚性的,僅在形成配子或隔離死菌絲的地方形成隔膜。 Zygomycota不再被認為是不被認為是真正單系的。
詞源
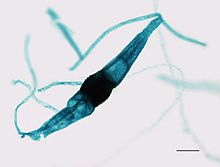
zygomycota的名稱是指由該進化枝的成員形成的Zygosporangia ,其中在有性繁殖過程中形成了抗性球形孢子。 Zygos是希臘語“加入”或“軛”,指的是兩個產生這些孢子的菌絲鏈的融合,而-mycota是指真菌分裂的後綴。
孢子

“孢子”一詞用於描述與傳播和擴散有關的結構。合子孢子可以通過性和無性含量形成。在發芽之前,孢子處於休眠狀態。在此期間,代謝率非常低,可能會持續幾個小時到很多年。休眠有兩種類型。外源休眠受環境因素(例如溫度或養分的可用性)控制。內源性或構型休眠取決於孢子本身的特徵。例如,代謝特徵。在這種類型的休眠狀態下,即使環境條件有利於生長,也可以預防發芽。
線孢子
在合子中,線孢子(孢子孢子孢子)是無性形成的。它們是由特殊結構(Sporangia)組成的,這些結構(Sporangia)含有數千到數千個孢子,具體取決於物種。 Mitosporangia由特種菌絲(孢子孢子菌)(孢子囊)攜帶。這些專門的菌絲通常表現出負重的引力和陽性光質主義,從而使孢子分散良好。孢子囊壁很薄,很容易被機械刺激(例如雨滴,通過動物)破壞,導致成熟的線孢子的擴散。這些孢子的壁含有某些物種中的孢子囊素。孢子囊素是由β-胡蘿蔔素形成的,對生物學和化學降解非常抗性。合子孢子也可以根據其持久性進行分類:
衣原體
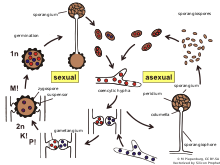
衣原體是無性孢子的,與孢子囊孢子不同。衣原體的主要功能是菌絲體的持久性,當菌絲體降解時它們會釋放。衣原體沒有分散的機制。在zygomycetes中,衣原體的形成通常是循環的。但是,它也可能是終端。根據其功能,衣原體有一個厚的細胞壁,並具有色素色。
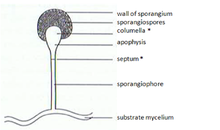
合子
合子是趨化的空中菌絲,是zygymycota的性器官,除了它們不是空中而是在底層中發現的植物科學。他們有兩種不同的交配類型(+)和( - )。相反的交配類型由於相反的鏈(主要是三孢子及其前體)的揮發性信息素而彼此相對。一旦兩種相反的交配類型建立了初始接觸,它們就會通過多個步驟產生zy孢子。
一旦建立了Zygophores之間的接觸,它們的牆壁彼此固定,將接觸位點變平,然後將接觸位點稱為融合隔隔。合子的尖端變得膨脹並形成所謂的progametangia。隔隔通過逐漸向內延伸而發展,直到將末端Gametangia與前提基鹼基分開為止。在這一點上,zygophore被稱為懸架。囊泡積聚在融合隔膜上,然後它開始溶解。在融合隔隔隔隔完全溶解之前的一小部分,主外壁開始變稠。隨著融合隔膜溶解,這可以看作是一壁上的深色斑塊。牆上的這些黑暗斑塊最終將發展成構成Zygospore牆厚度的疣結構。隨著zy孢子的擴大,疣狀結構也是如此,直到整個細胞周圍存在連續。在這一點上,電子顯微鏡無法再滲透壁。最終,疣穿過主要壁並變暗,這可能是黑色素引起的。
減數分裂通常發生在zy孢子發芽之前,並且有幾種可區分的核行為類型。 1型是在幾天內快速核融合的核融合的時候,導致具有單倍體核的成熟zygospore。 2型是在某些核不配對並退化的時候,減數分裂被延遲到發芽。 3型是單倍體核繼續通過有絲分裂分裂的,然後有些將一些相關分為組,而有些則沒有。這會導致在生殖孢子囊中發現二倍體和單倍體核。
細胞壁
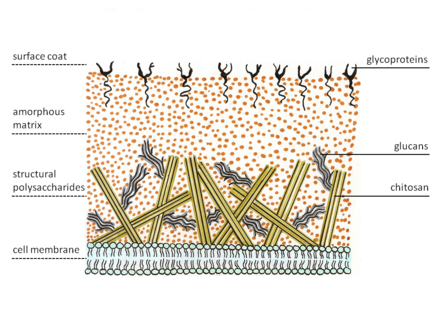
合子表現出細胞壁的特殊結構。大多數真菌具有幾背作為結構多醣,而zygomycetes合成了殼蛋白的脫乙酰化均聚物殼聚醣。幾丁質由β -1,4鍵合的N-乙酰葡萄糖胺構建。真菌菌絲在尖端生長。因此,專門的囊泡,殼聚醣將幾丁質的前體及其合成酶,幾丁質合成酶通過胞吐作用帶到膜的外部。膜上的酶從核苷酸糖底物,尿苷二磷酸- N-乙酰-D-葡萄糖胺催化糖苷鍵形成。然後,新生的多醣鏈被酶幾丁質脫乙酰基酶切割。該酶催化了幾丁質中N-乙酰胺基的水解裂解。之後,殼聚醣聚合物鏈形成微纖維。這些纖維嵌入由蛋白質,葡萄糖(推定的殼聚醣纖維交叉鏈接),甘露蛋白,脂質和其他化合物組成的無定形基質中。
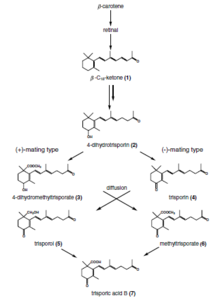
三孢子
Trisporic Acid是一種C-18萜類化合物,在合子菌中通過β-胡蘿蔔素和視黃醇途徑合成。它是負責真菌物種性別分化的信息素化合物。
歷史
1964年發現三孢子是一種代謝產物,導致Blakeslea trispora的胡蘿蔔素產生增強。後來證明是導致粘膜粘液群產生合子產生的激素。美國真菌學家和遺傳學家阿爾伯特·弗朗西斯·布萊克斯(Albert Francis Blakeslee)發現,某些粘膜是自我殺傷(異質化),其中兩種菌株(指定(+)和( - )的相互作用對於開始性活動是必不可少的。 Goettingen大學的Hans Burgeff發現這種相互作用是由於在底層和大氣中擴散的低分子量物質的交換。這項工作構成了任何真菌中性激素活性的首次演示。在粘液中闡明性相互作用的荷爾蒙控制延長了60年,涉及來自德國,意大利,荷蘭,英國和美國的真菌學家和生物化學家。
三孢子在粘液中的功能
對zygomycota中兼容的性伴侶的識別是基於三孢子酸的合作生物合成途徑。早期的三孢子衍生物和三孢子酸會誘導兩種潛在菌絲的腫脹,因此稱為合子,這些誘導劑分子的化學梯度會導致彼此的生長。這些progametangia彼此接觸並建立牢固的聯繫。在下一階段,建立隔膜是為了限制營養性菌絲體的發育中的zy孢子,並以此方式形成了伴侶形成吊帶和gametangia。溶解融合壁後,混合了兩個Gametangia的細胞質和大量核。選擇過程(未研究)導致核和減數分裂的減少(直到今天也沒有研究)。發生了幾種細胞壁修飾,以及孢子囊素的摻入(負責孢子的深色),從而導致成熟的同孢子。
作為這種識別途徑的終點,三孢子只能在兩個兼容伴侶的存在下產生,該伴侶在酶上產生三角孢子的前體,可以被潛在的性伴侶進一步利用。這些反應的物種特異性包括通過空間分離,衍生物的理化特徵(波動性和光敏度),三孢子蟲的化學修飾以及轉錄/轉錄後調節獲得的物種特異性。
寄生蟲主義
三孢子蟲也用於寄生蟲和宿主之間識別的介導。一個例子是觀察到寄生蟲寄生蟲的寄生蟲性質的宿主 - 寄生蟲相互作用,寄生蟲是Zygomycetes的輔助分枝桿菌和Absidia Glauca 。這種相互作用是生物營養融合寄生蟲主義的一個例子,因為遺傳信息被轉移到宿主中。與zy孢子形成相比,許多形態學相似性都可以看到,但是成熟的孢子稱為錫克伊索孢子,是寄生的。在此過程中,gall樣結構由宿主的吸毒glauca產生。這與進一步的證據相結合,導致假設三孢子蟲不是嚴格的物種特異性,而是它們可能代表粘膜交配識別的一般原則。
光質主義
在zygomycetes phycomyces blakesleeanus , Mucor Circinelloides和Pilobolus Crystallinus中已經研究了光調節。例如,在Pilobolus crystallinus中,光負責分散機理,而植物科動物的孢子囊孢子菌的孢子體向光生長。當光,尤其是藍光參與真菌發育的調節時,它會指導真菌結構的生長並激活代謝途徑。例如,zygomycota使用光作為信號來促進營養繁殖和空中菌絲的生長以促進孢子分散。
真菌光質主義已通過植物菌作為模型的孢子蟲孢子體進行了詳細研究。 Phycomyces具有復雜的光感受器系統。它能夠對不同的光強度和不同的波長反應。與對藍光的積極反應相反,對紫外線也有負反應。還觀察到對紅光的反應。
通過光激活β-胡蘿蔔素生物合成
植物去飽和酶(碳水化合物)和雙功能性植物合酶/胡蘿蔔素環烯酸酶(植物科中的CARRA,粘膜中的CARRA)的兩個基因負責合成β-胡蘿蔔素。在粘液中發現的基因CRGA的乘積通過抑制碳水化合物和Carrp mRNA的積累來抑制胡蘿蔔素的形成。
光在孢子形成和性發展中的影響
Zygomycete P. blakesleeanus構建了兩種類型的孢子囊,巨摩托車和大小不同的顯微鏡。這些孢子囊的形成在不同的光線上起作用,因此在特定的感光體上起作用。光還調節無性孢子形成。在粘膜中,CRGA基因的產物充當激活劑。相反,由於專門的感光體系統,光抑制了植物科學的性發育。
引力症
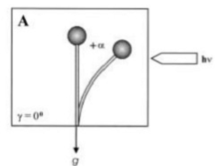
重力主義是植物或真菌響應重力的轉向或生長運動。這在兩個王國中同樣普遍。在真菌和植物中都需要史地那石,以實現重力感應的機理。 Zygomycota孢子囊源自專門的“基底菌絲”,並經過幾個獨特的發育階段,直到釋放出成熟的無性孢子。除了陽性光療法之外,孢子囊還通過負重壓力反應指向適合孢子分散和分佈的位置。這兩種反應都是生長反應,即彎曲是由於孢子體的相對側面的不同生長引起的,並相互影響。植物菌引力反應機理的唯一模型是基於周圍細胞質內液泡的浮動性。提出了所得的細胞質的不對稱分佈,以產生地平線放置孢子囊下部的壁生長增加,就像在形成較厚的較厚的細胞質層中,分泌細胞壁材料的囊泡數量會高於上側。大約15 - 30分鐘的水平放置的孢子蟲生物團在大約15-30分鐘後開始,一直持續到大約12-14小時後,孢子囊尖端已恢復了其原始的垂直位置。通常,與光營養的反應相比,重力反應較弱。但是,在某些條件下,可以建立平衡,並且反應是可比的。在植物和真菌中,光質主義和重力主義以復雜的方式相互作用。在單側光線連續輻照過程中,zygomycete真菌的孢子蟲(果實體),植物科動物的blakesleeanus達到了光膜片平衡的彎曲角,在該平衡中,引力和光子刺激彼此之間平衡了重力和光子刺激(圖1,彎曲角度+α,到來的角度+α)輻照)。

涉及增壓劑的蛋白質晶體
在植物科動物中,野生型孢子囊包含易於看到的八面體paracrystalline晶體,尺寸高達5×5×5μm。通常,它們在由十多個晶體組成的簇中的主要液泡附近發現。它們通常與液泡的轉讓。當孢子團傾斜時,可以觀察到以約100μm/s的速度沉積。在沉積過程中滑動或在液泡膜上拉動和transepts作為可能的細胞骨架反應,並激活位於細胞膜中的受體。這些受體反過來觸發了一系列事件,最終導致細胞壁的不對稱生長。對野生型和突變菌株孢子體生長的彎曲角的研究表明,沒有晶體的突變菌株表現出降低的降壓反應。
脂肪液滴
頂端脂質球的複合物也參與了顆粒受感受。這些脂質聚集在細胞結構中,脂質球的複合物,高於頂端的尖端約0.1mm。 (圖2)當孢子蟲形成時,球遷移到小柱。在成熟的階段,由於其浮動性,這種複合物被認為是重量受體。缺乏這種脂質複合物的突變體顯示大大降低了重力反應。
系統發育
從歷史上看,所有產生zy孢子的真菌都被認為是相關的,並將其置於zygomycota中。分子系統發育學的使用越來越多地揭示了這種分組是銷子的。但是,這些進化枝的等級(即,門或亞球)存在爭議。接下來是真菌的系統發育,其Zygomycete subphyla源自Spatafora等。 (2016年)都有可能的門名。
| |||||||||||||||||||
| |||||||||||||||||||
工業用途
許多合子菌可以用於重要的工業過程中。桌子中介紹了它們的簡歷。
| 物種 | 產品 | 用途 |
|---|---|---|
| 幾種粘液和根莖屬。 | 脂肪酶和蛋白酶 | 皮革,洗滌劑和醫療行業(類固醇轉化) |
| 根莖 | 纖維酶 | 食品生產(即,坦佩) |
| R. Oryzae ,其他根莖屬。 | 富馬酸 | 各種各樣的 |
| Rhizopus spp。 | 乳酸 | 各種各樣的 |
| R. Delemar | 生物素 | 各種各樣的 |
| Mortierella Romanniana , Mortierella vinacea和Mucor Insidus | 亞麻酸 | 各種各樣的 |
| Mortierella alpina | 花生四烯酸 | 各種各樣的 |
| Blakeslea Trispora | β-胡蘿蔔素 | 各種各樣的 |
培養條件

合子能夠在各種環境中生長。它們中的大多數是嗜嗜性的(在10-40°C下生長為最佳20–35°C),但是有些人(例如粘膜miehei或粘膜pusillus )是嗜熱的,最小生長溫度約為20°C,最大延伸至最高延伸至60°C。其他類似Mucor Hiemalis的人可以在低於0°C的溫度下生長。
粘膜的某些物種能夠在厭氧條件下生長,而大多數則需要有氧條件。此外,雖然大多數合子菌僅在高水活性下生長,但其中一些能夠以至少15%的鹽濃度生長。大多數粘液在室溫下在瓊脂上迅速生長,並在2-3天內用粗大的空中菌絲體填充培養皿。當在半動植物條件下在液體培養中孵育時,幾種物種像州一樣在酵母中生長。在較高的孵育溫度(30-40°C)下,可以刺激zy孢子形成。

固體瓊脂中zygomycota的生長會產生低或非常高的纖維菌落,從而迅速填充整個培養皿。它的顏色可能從純白色到灰色陰影。在舊文化中,觀察到深色色素的孢子囊。一切都取決於物種和所使用的媒體。在液體培養中,zygomycota通常形成平淡的質量,不會產生孢子。這是因為它們不能生長空中菌絲。
文化傳媒
合子在大多數標準的真菌培養基(例如Sabouraud Dextrose瓊脂)上生長良好。它們也可以在選擇性和非選擇性媒體上成長。也可以使用最少的媒體,補充媒體和感應媒體。大多數合子對環己酰亞胺(Actidione)敏感,該試劑不應用於培養基中。
再生產
zygomycete的一個常見例子是黑麵包黴菌( Rhizopus Stolonifer ),這是粘膜的成員。它散佈在麵包和其他食物來源的表面上,使菌絲向內吸收營養。在其無性階段,它在直立菌絲的尖端上形成球莖黑色孢子囊,每個孢子均包含數百個單倍體孢子。
與大多數zygomycetes一樣,無性繁殖是最常見的繁殖形式。與其他zygomycetes一樣,當不同交配類型的單倍體菌絲彼此彼此接近時,就會發生在根瘤菌中的有性繁殖。 Gametangia的生長開始接觸後開始,漿細胞或細胞質的融合發生。核納米是核的融合,緊隨其後。然後是Zygosporangia 。 Zygosporangia通常是厚壁的,對環境困難高,並且代謝惰性。但是,當條件改善時,它們會發芽以產生孢子囊或營養菌絲。減數分裂發生在Zygosporangium的發芽期間,因此產生的孢子或菌絲是單倍體。在溫暖和潮濕的條件下生長。
一些合子菌以更精確的方式分散其孢子,而不是簡單地允許它們在氣流上漫無目的地漂移。 Pilobolus是一種在動物糞便上生長的真菌,借助輕敏感的色素( β-胡蘿蔔素)將其孢子囊彎曲朝向光,然後用高壓細胞質的爆炸性噴射來“發射”它們。 Sporangia可以發射至2 m,將它們遠離糞便遠處,並希望將其植物放在植被上,該植被將被草食動物食用,最終將其存放在其他地方的糞便中。扎氏菌群昆蟲侵蝕性的成員之間已經進化了刺激孢子排放的不同機制。
分生孢子的進化
分生孢子從孢子孢子的演變是zygomycetes和ascomycetes之間的主要定義差異。合子菌對分生孢子的孢子孢子的演變類似於在comcycetes中發現的分生孢子,可以通過在zygomycetes中看到的一系列形式來建模。許多合子菌在單個孢子囊內產生多個孢子囊。有些進化出了多個含有孢子囊孢子的小孢子囊。在某些情況下,每個孢子囊中可能有三個孢子,而幾個物種的孢子囊僅包含一個孢子。 Zygomycete的Choanephora具有一個孢子囊,其中包含一個帶有孢子壁的孢子,在孢子囊底部可見。該結構類似於一個分生孢子,該分生孢子有兩個融合的細胞壁,一個內孢子壁和一個外孢子壁。