借助胎兒
| 借助胎兒 時間範圍:巴雷米亞人, | |
|---|---|
 | |
| 用手重建生活 | |
| 科學分類 | |
| 王國: | Animalia |
| 門: | Chordata |
| 進化枝: | 恐龍 |
| 進化枝: | Saurischia |
| 進化枝: | Theropoda |
| 家庭: | †Trodondidae |
| 屬: | †借助胎兒 沉等。,2017年 |
| 類型物種 | |
| †借助貸方Curriei 沉等。,2017年 | |
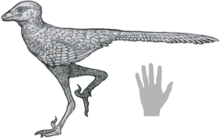
借助胎兒(意義 ”騙子獵人”)是屬的TrodontidTheropod恐龍來自早期白堊紀的中國。它包含一個物種,L. Curriei, 而得名古生物學家菲利普·庫裡(Phillip J. Currie)2017年,Shen Cai-Zhi及其同事來自一個明顯的,幾乎完整的骨骼,這是最完整的troodindid標本之一。沉及其同事發現了放置的指示性狀借助胎兒在Trodondidae內部。這些特徵包括其眾多,小且緊密的牙齒,以及尾巴末端的椎骨,具有淺凹槽代替神經刺在他們的頂表面。
在Trodondidae內部,最接近的親戚借助胎兒曾是eosinopteryx,它也與錨和小米;儘管這些傳統上被放置在Trodondidae外面,但系統發育分析由Shen及其同事進行的證據支持這些證據帕拉維亞人作為troodontids。但是,與其近親相比借助胎兒腿相對較長,尤其是股骨。作為從地理和時間上可比的第五個troodindid地層,借助胎兒增加了中國多齒的多樣性。
描述
總體長度(估計的頭骨,頸部,軀乾和尾巴的長度)約為69厘米(27英寸)借助胎兒很小Trodontid.[1]
頭骨和下巴
借助胎兒具有長,狹窄和三角形的頭骨,長9.8厘米(3.9英寸)。在鼻子的前面,喜歡錫諾芬劑和xixiasaurus, 這前甲蟲排除上頜骨從鼻孔的邊緣。上頜骨的表面有三個開口,上頜骨,上頜和防肌fenestrae。下面,上頜骨形成次生口味如Byronosaurus,Gobivenator, 和xixiasaurus。在troodindids中獨特,牙本質骨苗條並輻射成三個過程。喜歡Zanabazar,有一個氣動憩室在裡面jugal骨骨頭內存在空氣囊;後面還有一個氣動開口四邊形骨,就像其他troodontids一樣。[2]與眾不同Sauronithoides,Zanabazar, 和Stenonychosaurus,劃分的波峰頂骨不參加頭骨後部的顳上葉氏(上)顳葉的邊界。[1]
troodontids的特徵,[2]借助胎兒在其淺和三角形下頜的外邊緣上有一個凹槽的凹槽。下頜的底部邊緣略微凸出;在Sinornithoides,這是筆直的。這牙齒和角骨可能已經形成了下頜內的柔性接頭 - 即,輔助接頭。[3]與眾不同小米,牙齒和上頜骨在下顎的同一位置終止。也像其他troodontids(有鼻竇例外),借助胎兒有許多小的,緊密的牙齒,上顎至少有15個,下頜23。與許多人不同基礎Trodontids。上頜骨的牙齒行終止於孔孔式芬斯特拉的前緣以下,而它則進一步向前終止 - 上頜fenestra的後部 - Jinfengopteryx.[1]
椎骨
有十個頸椎,十二背(軀幹)椎骨,至少有16個尾骨(尾巴)椎骨借助胎兒。在頸椎中,第三到第八的伸長,第五是最長的。之間關節過程第五宮頸腔也被稱為前卵形植物,也有最長的時間。在第三和第四個宮頸中,後者像其他宮頸衍生的troodontids,前雜誌植物會長於另一組稱為Zygapophyses的過程。在背側,相對於錨。在尾巴中,過渡點 - 尾部側面變得更加壓縮的點,使它們是亞三角形而不是矩形 - 發生在第七尾部,而不是向前Sinornithoides和梅(在第九次出現的地方)。[4][5]最長的尾是第十四個,幾乎是第六次長度的兩倍。在尾部的底面人字形略微彎曲和向後定向,如Deinonychus.[1]
四肢
這Acromion過程肩cap的借助胎兒與基礎troodontids一樣,發育不良。但是,與基底troodontids不同,關節盂 - 手臂插座 - 垂直向下而不是側向側面。在肱骨,三角洲冠延長了骨頭長度的40%,並終止了骨頭的一半。肱骨短,相對於股骨, 在借助胎兒(長度為59%)eosinopteryx(長度為80%)。[6]在下面,三位數的手是獨一無二的方陣第一個數字比第二位數長metacarpal,是後者長度的1.49倍。[1]
如梅,頂部ilium具有彎曲的正弦形狀借助胎兒。上面沒有波峰actabulum(臀部插座)的借助胎兒,不同錨。iLium的後部(後移動後過程)的底部邊緣也比兩者都短錨和eosinopteryx。獨特的是,在頂端的頂端沒有任何過程坐姿,骨頭也有細長閉孔過程。這恥骨指向向前借助胎兒,但倒退梅.[5]此外,借助胎兒是軀幹長度的兩倍,而梅它是軀幹長度的2.8倍。與眾不同錫諾芬劑,借助胎兒有個第四騎手在股骨上。這脛骨是細長的,是股骨長度的1.4倍Sinornithoides。四位數的腳高度緊湊,有瞼板那狹窄到底部。第三metatarsal偏離第二和第四,在後兩者之間形成一個比其他trodondids更深的槽。按比例,第二位數的第一個指標相對於第二個卵形較短借助胎兒(長135%)錫諾芬劑(長度為150%)。[1][7]
發現和命名
借助胎兒從單個標本中得知,這是一個幾乎完整且保存完好的骨骼,其原始鉸接位置保留了大多數骨骼。它在盧吉亞頓床Yixian組在上海比比亞,騙子,中國;目前,它存儲在達利安自然歷史博物館(DNHM)達利安,在加入編號DNHM D3012下進行借用。一些標本的鼻骨骨頭不完整,非法化石商人增加了一些腳趾骨;因此,它是有史以來發現的最完整的Trodontid化石之一。它的頭向前彎曲,四肢被塞進去。這與經典不同死亡姿勢(頭部向後彎曲的地方)以及睡眠的姿勢梅和Sinornithoides.[1]
2017年,DNHM D3012被任命為類型標本新屬和物種借助貸方Curriei由Shen Caizhi,Zhao Bo,Gao Chunling,LüJunchang和MartinKundrát。屬名稱借助胎兒結合後綴-venator,意思是“獵人”拉丁;特定名稱Curriei尊重加拿大古生物學家菲利普·約翰·柯里(Philip John Currie)研究小獸腳類動物。[1]
分類
Shen及其同事確定借助胎兒作為Trodondidae的成員,基於其眾多的,緊密間隔的牙齒,這些牙齒被限制在下方王冠;其四邊形後部的氣動開口;橢圓形的形狀大孔;替換神經刺通過椎骨的淺中線凹槽,尾巴的末端;高度上升的過程阿斯托拉拉斯;及其不對稱和亞容量測量(即,第三個meta骨有點被相鄰的meta骨捏住)腳。[2][8]他們進一步將其放在“較高的troodontid進化枝”中,基於缺乏球形膠囊狀結構偏盤它的口感,以及promaxillary fenestra在其頭骨上的存在。[1]
基於a系統發育分析從Takanobu Tsuihiji及其同事在2016年進行的先前分析進行了修改,而Gao及其同事在2012年對這一修改進行了修改。XU Xing和同事在2012年[9]沉及其同事發現借助胎兒組成一個統一的小組,或進化枝, 和eosinopteryx,錨, 和小米,因此為這些傳統放置提供了相反的證據分類單元作為非troodontid成員paraves。它們是由於牙齒被扁平和彎曲而團結的,牙齒在牙齒行的中間的高度小於其寬度的兩倍。Acromion的前邊緣被遠足了;雅親子和類囊樣;在手指上的第二位數字的第一個緣彈上存在法蘭;和向後的恥骨。[1]
在這個進化枝中,沉及其同事沒有命名,借助胎兒與eosinopteryx儘管錨與小米。前兩個是由於下頜前部缺乏鋸齒的牙齒而團結的。頭骨超過股骨長度的90%以上;具有細長軸的頸肋骨比相應的椎骨更長。ilium的前端是“輕柔的”。同時,後兩個人在背面的扇形上分享了神經刺的尖端。冠狀球體是三角形的;手的第一個數字上的爪子被強烈拱起,高於鉸接表面的頂部。第二和第三位數在爪子的頂端存在“唇”;恥骨軸的前邊緣是凸面;腳上的第三和第四位爪子強烈彎曲。系統發育分析的結果在以下系統發育樹.[1]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
組織學
薄部分脛骨的整型標本借助胎兒表示皮質骨為1.5毫米(0.059英寸)厚。皮質被分為四個區域被捕生長線(滯後),這表明該動物死亡時至少四歲(進一步的滯後可能因擴張而消除髓腔)。Shen及其同事根據最內向區的薄度與2區相比,它已部分侵蝕。但是,完整的區域甚至更薄,僅是區域2寬度的四分之一,而區域4甚至更薄(儘管不完整)。這表明到第三年末,增長已經大大放緩。[1]
每個滯後都被兩個緻密的血管造成的骨骼(即缺乏血管開口)圍繞,而沉及其同事稱為“預註冊”和“後”和“後”。第一個滯後與其他滯後不同,因為它由兩個滯後組成,一個滯後比另一個較弱,表明生長在恢復骨骼生長之前會輕微放慢。骨外側邊緣上沒有外部基本系統(EFS),表明整型在死亡時仍在增長。然而,區域的薄度降低,外層層中血管骨的存在以及證據骨頭重塑共同表明它接近骨骼成熟。[1]
古生態學
根據沉的同事的說法借助胎兒是從中國聞名的11個troodontids之一,也是第五個白堊紀的中國troodontid錫諾芬劑,鼻竇,梅, 和Jinfengopteryx.[1]在2017年的單獨出版物中,Shen也是主要作者,並描述了另外一部Troodontid,達利亞龍,哪個形成Sinovenatorinae與前三個。[10]除了Jinfengopteryx,所有這些Troodontids都住在Lujiatun床上。而沉和同事將Lujiatun床分配給HauterivianChang Su-Chin及其同事發布的較新的日期估計值,這是〜126歲的年齡段嘛對於Lujiatun床,可追溯到巴雷米亞人時代。[11]
同時的恐龍包括微療法龍龍Gracilaptor;這Oviraptorosaur牙龍;這Ornithomimosaurs深圳[12]和十六進制;[13]這前龍霸王龍迪隆;[12]這鈦龍形蜥腳類動物Euhelopus;[14]這OrnithopodJeholosaurus;和Ceratopsians例如無處不在Psittacosaurus[15]也Liaoceratops.[12]包括哺乳動物貴族,[16]Gobiconodon,juchilestes,Maotherium,Meemannodon, 和repenomamus.[17][18][19]其他四腳架包括青蛙Liaobatrachus[20]和腸子;[21]和蜥蜴Dalinghosaurus.[22]Lujiatun床包括河流和火山碎裂沉積物,表明有火山的河流景觀,[1]可能通過拉哈爾.[19]該區域的平均每年氣溫至少達到10°C(50°F)。[23]
也可以看看
參考
- ^一個bcdefghijklmnoShen,C.-Z。趙,b。Gao,C.-L。;Lu,J.-C。;Kundrát,M。“新的Trodontid恐龍(借助貸方CurrieiGen。et sp。nov。)從西鋰省的白堊紀早期的伊克斯島形成”。Acta Geoscientica Sinica.38(3):359–371。doi:10.3975/cagsb.2017.03.06.
- ^一個bcMakovicky,P.J。; Norell,M.A。(2004)。“ trodondidae”。在Weishampel,D.B。;Dodson,P。;Osmólska,H。(編輯)。恐龍(第二版)。伯克利:加利福尼亞大學出版社。第184-195頁。
- ^Tsuihiji,T。;Barsbold,R。;M. Watabe;Tsogtbaatar,K。;Chinzorig,T。;富士山;Suzuki,S。(2014)。“一種精美的troodontid heropod,擁有有關蒙古上白堊統的新信息的新信息”。NaturWissenschaften.101(2):131–142。Bibcode:2014nw .... 101..131T.doi:10.1007/S00114-014-1143-9.PMID 24441791.S2CID 13920021.
- ^Currie,P.J。;Dong,Z。(2001)。“來自中華人民共和國的白堊紀巨豆植物(恐龍)的新信息”。加拿大地球科學雜誌.38(12):1753–1766。doi:10.1139/e01-065.
- ^一個bGao,C。;E.M. Morschhauser;Varricchio,D.J。;Liu,J。;Zhao,B。(2012)。“第二個睡眠的龍:中國troodontid的新解剖細節梅·朗(Mei Long)對系統發育和taphonomy的影響”.PLOS ONE.7(9):E45203。doi:10.1371/journal.pone.0045203.PMC 3459897.PMID 23028847.
- ^Godefroit,p。;Demuynck,H。;Dyke,G。;胡,d。;Escuillié,F.O。Claeys,P。(2013年)。“降低了新的侏羅紀帕拉維亞獸腳類腳類犬的羽毛和飛行能力”.自然通訊.4:1394。Bibcode:2013NATCO ... 4.1394G.doi:10.1038/ncomms2389.PMID 23340434.
- ^Xu,X。;Zhao,ji;沙利文,c。Tan,Q.-W。;桑德(M。)MA,Q.-Y.(2012)。“ Trodondid IVPP V 10597的分類法重新考慮”(PDF).椎骨palasiatica.50(2):140–150。
- ^特納,A.H。;Makovicky,P.J。;Norell,M.A。(2012)。“對龍龍系統學和帕拉維亞系統發育的綜述”.美國自然歷史博物館的公告.371:1–206。doi:10.1206/748.1.HDL:2246/6352.S2CID 83572446.
- ^Tsuihiji,T。;Barsbold,R。;M. Watabe;Tsogtbaatar,K。;S.S. Suzuki;Hattori,S。(2016)。“蒙古下白堊統的trodondid theropod(恐龍:Saurischia)的新材料”。歷史生物學.28(1-2):128–138。doi:10.1080/08912963.2015.1005086.S2CID 128725436.
- ^Shen,C。;呂,J。;Liu,s。;Kundrát,M。;Brusatte,S.L。;Gao,H。(2017)。“來自中國獅子省的下白堊統伊克斯島形成的新的troodontid恐龍”(PDF).Acta Geologica Sinica.91(3):763–780。doi:10.1111/1755-6724.13307.
- ^Chang,S.-C。;Gao,K.-Q。;週,Z.-F。;Jourdan,F。(2017)。“對Yixian形成的新的地層學約束,對Jehol Biota產生了影響”。古地理,古氣候學,古生態學.487:399–406。Bibcode:2017ppp ... 487..399c.doi:10.1016/j.palaeo.2017.09.026.
- ^一個bcXu,X。; Norell,M.A。(2006)。“來自中國西部的白堊紀耶霍爾集團的非阿維亞恐龍化石”(PDF).地質雜誌.41(3–4):419–437。doi:10.1002/gj.1044。存檔原本的(PDF)在2016-03-03。檢索2017-12-03.
- ^Jin,L。; Jun,c。; Godefroit,P。(2012)。“新的基底鳥瘤(恐龍:theropoda)來自中國東北部的白堊紀伊克斯島形成”(PDF)。在Godefroit,P。(編輯)。Bernissart恐龍和早期白堊紀陸地生態系統。過去的生活。布盧明頓:印第安納大學出版社。pp。467–487。ISBN 978-0-253-35721-2.
- ^Barrett,P.M。;王,X.-L。(2007)。“基底鈦龍(恐龍,蜥蜴龍)牙齒,來自中國富裕省的下白堊紀伊克斯島形成”。古羅德.16(4):265–271。doi:10.1016/j.palwor.2007.07.001.
- ^赫德里克(B.P。); Dodson,P。(2013年)。“ Lujiatun psittacosaurids:使用3D幾何形態計量學了解個體和Taphonomic變異”.PLOS ONE.8(8):E69265。Bibcode:2013 Ploso ... 869265H.doi:10.1371/journal.pone.0069265.PMC 3739782.PMID 23950887.
- ^胡,y。Meng,J。;李,c。;Wang,Y。(2010)。“來自中國獅林早期白堊紀耶霍爾生物群的新基底哺乳動物”.皇家學會b會議錄.277(1679):229–236。doi:10.1098/RSPB.2009.0203.PMC 2842663.PMID 19419990.
- ^Wang,Y.-Q。;Hu,Y.-M。;Li,C.-K。(2006)。“中國中生代哺乳動物研究的最新進展綜述”(PDF).椎骨palasiatica.44(2):193–204。
- ^Lopatin,A。; Averianov,A。(2015)。 “Gobiconodon(哺乳動物)來自蒙古早期的白堊紀和戈比科尼迪科的修訂”。哺乳動物進化雜誌.22(1):17–43。doi:10.1007/s10914-014-9267-4.S2CID 18318649.
- ^一個b江,b。fürsich,f.t。;Sha,J。;王,b。Niu,Y。(2011)。“中國西部的西部胎兒的早白堊統火山及其對化石保存的影響”。古地理,古氣候學,古生態學.302(3):255–269。doi:10.1016/j.palaeo.2011.01.016.
- ^Dong,L。;Roček,Z。;王,y。瓊斯,M.E.H。(2013)。“來自中國西部獅子的下白堊統耶霍爾集團的阿努拉人”.PLOS ONE.8(7):E69723。doi:10.1371/journal.pone.0069723.PMC 3724893.PMID 23922783.
- ^王,y。瓊斯,M.E.H。;埃文斯(S.E.)(2007)。“下白堊統的jiufotang組的少年阿努蘭,中國騙子”(PDF).白堊紀研究.28(2):235–244。doi:10.1016/j.cretres.2006.07.003。存檔原本的(PDF)在2017-10-25。檢索2017-12-03.
- ^埃文斯(S.E.);王,y。瓊斯,M.E.H。(2007)。“來自中國下白堊紀的蜥蜴骨骼的聚集”。Senckenbergiana Lethaea.87(1):109–118。doi:10.1007/bf03043910.S2CID 83907519.
- ^Amiot,R。;Wang,X。;週,Z。;Wang,X。;Buffetaut,E。;Lécuyer,c。;ding,z。;Fluteau,F。;Hibino,T。;Kusuhashi,n。Mo,J。;Suteethorn,V。;王,y。Xu,X。;Zhang,F。(2011)。“東亞恐龍的氧同位素揭示出異常寒冷的白堊紀氣候”.美國國家科學院論文集.108(13):5179–5183。doi:10.1073/pnas.1011369108.PMC 3069172.PMID 21393569.
















