進化
進化是連續幾代人生物種群的遺傳特徵的變化。當諸如自然選擇和遺傳漂移之類的進化過程對遺傳變異作用時,就會發生進化,從而導致某些特徵或多或少在連續幾代人的人群中變得越來越普遍。進化的過程在每個級別的生物組織中都引起了生物多樣性。
自然選擇的進化論是由查爾斯·達爾文(Charles Darwin)和阿爾弗雷德·羅素·華萊士(Alfred Russel Wallace)在19世紀獨立構想的,以解釋為什么生物適應其物理和生物學環境。該理論首先是在達爾文的《物種起源》的書中詳細介紹的。關於生物體的可觀察到的事實可以確定自然選擇的進化:(1)通常產生更多的後代,而不是生存; (2)特徵在個人的形態,生理和行為方面有所不同; (3)不同的特徵賦予不同生存和繁殖率(差異適應性); (4)特徵可以世代相傳(健身的遺傳力)。因此,連續幾代人,人口的成員更有可能被對該環境有利的父母的後代所取代。
在20世紀初期,駁斥了進化的競爭觀念,並將進化與門德爾的遺傳和人口遺傳學相結合,從而引起現代進化論。在這種綜合中,遺傳的基礎是在DNA分子中產生的一代信息。改變人群中DNA的過程包括自然選擇,遺傳漂移,突變和基因流動。
地球上的所有生命(包括人類)都展示了最後一個普遍的共同祖先(Luca),該祖先大約在3.5億年前。化石記錄包括從早期生物石墨到微生物墊化石再到化石多細胞生物的進展。現有的生物多樣性模式已通過新物種的重複形成(物種),物種內部的變化(異性)以及物種(滅絕)(滅絕)的反复形成形成。在共有一個較新的共同祖先的物種中,形態學和生化特徵往往更相似,這些物種歷史上被用來重建系統發育樹,儘管遺傳序列的直接比較是當今更常見的方法。
進化生物學家繼續通過形成和檢驗假設來研究進化的各個方面,並根據現場或實驗室的證據以及數學和理論生物學方法產生的數據來構建理論。他們的發現不僅影響了生物學的發展,還影響了包括農業,醫學和計算機科學在內的其他領域。
遺傳

生物體的進化是通過遺傳特徵的變化(生物的遺傳特徵)發生的。例如,在人類中,眼睛的顏色是一種遺傳的特徵,一個人可能會從他們的一位父母那裡繼承“棕眼特徵”。遺傳性狀由基因控制,並且在生物體基因組(遺傳物質)中的完整基因稱為其基因型。
組成生物體的結構和行為的一組可觀察的特徵稱為其表型。其中一些特徵來自其基因型與環境的相互作用,而其他特徵則是中性的。某些可觀察的特徵沒有繼承。例如,曬黑的皮膚來自一個人的基因型和陽光之間的相互作用。因此,曬黑不會傳遞給人們的孩子。表型是暴露於陽光時皮膚棕褐色的能力。但是,由於基因型變異的差異,有些人比其他人更容易曬黑。一個明顯的例子是具有遺傳性白化病特徵的人,他們根本不曬黑並且對曬傷非常敏感。
可遺傳的特徵通過DNA從一代傳遞到第二代,這是一種編碼遺傳信息的分子。 DNA是由四種類型的鹼基組成的長生物聚合物。沿特定DNA分子的鹼基序列以類似於拼寫句子的字母序列的方式指定了遺傳信息。在細胞分裂之前,複製了DNA,因此所得的兩個細胞中的每個細胞都會繼承DNA序列。指定單個功能單元的DNA分子的部分稱為基因。不同的基因具有不同的鹼基序列。在細胞內,每根DNA的長鏈稱為染色體。染色體中DNA序列的特定位置稱為基因座。如果一個基因座的DNA序列在個體之間有所不同,則該序列的不同形式稱為等位基因。 DNA序列可以通過突變改變,產生新等位基因。如果一個基因內發生突變,則新等位基因可能會影響基因控制的特徵,從而改變生物體的表型。但是,儘管在某些情況下,等位基因和特徵之間的這種簡單對應關係起作用,但大多數特徵都以定量或上位的方式影響了多個基因。
變化來源


如果人群內部存在遺傳變異,可能會發生進化。變異來自基因組突變,通過有性繁殖對基因改組以及種群之間的遷移(基因流)。儘管通過突變和基因流不斷地引入新的變異,但該物種的所有個體的大多數基因組都非常相似。但是,進化發育生物學領域的發現表明,即使基因型的差異相對較小,也會導致物種內部和物種之間的表型差異。
單個生物體的表型既來自其基因型和它所生活的環境的影響。現代進化合成將進化定義為這種遺傳變異的變化。相對於該基因的其他形式,一個特定等位基因的頻率或多或少會變得普遍。當新等位基因達到固定點時,變化就會消失 - 當它從人口中消失或完全取代祖先等位基因時。
突變
突變是細胞基因組的DNA序列的變化,是所有生物體遺傳變異的最終來源。當突變發生時,它們可能會改變基因的乘積,或防止基因發揮作用或沒有作用。
蛋白質編碼基因編碼區域中約有一半的突變是有害的 - 另一半是中性的。該地區總突變的一小部分賦予了健身益處。基因組其他部分的某些突變是有害的,但絕大多數是中性的。一些是有益的。
突變可能涉及染色體複製的大部分(通常是通過遺傳重組),該染色體可以將基因的額外副本引入基因組中。基因的額外副本是新基因進化所需的原材料的主要來源。這很重要,因為大多數新基因都會從共同祖先的先前存在的基因中演變出來。例如,人的眼睛使用四個基因使有意義的結構:三個用於彩色視覺,一個用於夜視;這四個是從單個祖先基因中降低的。
當重複的拷貝突變並獲取新功能時,可以從祖先基因產生新基因。一旦基因重複,此過程就更容易,因為它增加了系統的冗餘。一對中的一個基因可以獲取新功能,而另一個副本繼續執行其原始功能。其他類型的突變甚至可以從以前非編碼的DNA產生全新的基因,這種現象稱為“從頭基因出生” 。
新基因的產生也可能涉及復制幾個基因的小部分,然後將這些片段重新組合以形成新功能(外顯子洗牌)的新組合。當新的基因通過改組的預先存在的部分組裝而成時,域充當具有簡單獨立函數的模塊,可以將其混合在一起以產生具有新和復雜功能的新組合。例如,聚酮化合物合酶是製造抗生素的大酶。它們包含多達100個獨立的域,每個域都會在整個過程中催化一個步驟,就像彙編線中的一步一樣。
突變的一個例子是野豬小豬。它們是偽裝的,並顯示了深色和輕度縱向條紋的特徵模式。但是,黑色素皮質素1受體( MC1R )中的突變破壞了模式。大多數豬品種都帶有MC1R突變,破壞了野生型顏色和不同的突變,導致占主導地位的黑色著色。
性與重組
在無性生物體中,基因被遺傳在一起或鏈接,因為它們在繁殖過程中無法與其他生物的基因混合。相比之下,性生物的後代包含通過獨立分類產生的父母染色體的隨機混合物。在稱為同源重組的相關過程中,性生物在兩個匹配的染色體之間交換DNA。重組和重組不會改變等位基因的頻率,而是改變等位基因相互關聯的相互關聯,從而與等位基因的新組合產生後代。性別通常會增加遺傳變異,並可能增加進化速度。

性交的成本是約翰·梅納德·史密斯(John Maynard Smith)首先描述的。第一個成本是,在性二態性的物種中,只有兩個性別中的一個可以年輕。這種成本不適用於大多數植物和許多無脊椎動物的雌雄同體。第二個成本是,任何重現性行為的人只能將其50%的基因傳遞給任何個人後代,而隨著每一代的流逝,任何人的傳球都更少。然而,有性繁殖是真核生物和多細胞生物之間繁殖的更常見的手段。紅皇后假設已被用來解釋有性繁殖的意義,以此作為在不斷變化的環境中與其他物種進行進化的持續進化和適應性的手段。另一個假設是,性繁殖主要是促進生殖線DNA損傷的準確重組修復的適應性,而增加的多樣性是該過程的副產品,有時可能是適應性的。
基因流
基因流量是種群之間和物種之間的基因的交換。因此,它可以是人口新的變異來源或物種。基因流量可能是由個體之間的個體在生物體之間的運動引起的,這可能是由於內陸和沿海種群之間的小鼠運動可能引起的,或者在重金屬耐受和重量敏感人群之間的花粉運動的運動可能引起。草。
物種之間的基因轉移包括雜交生物的形成和水平基因轉移。水平基因轉移是遺傳物質從一種生物體轉移到另一生物的,而不是其後代。這在細菌中最常見。在醫學中,這有助於抗生素耐藥性的擴散,因為當一種細菌獲得抗性基因時,它可以迅速將其轉移到其他物種上。已經發生了從細菌到真核生物的水平轉移,例如釀酒酵母和Adzuki豆象鼻蟲Callosobruchus chinensis 。大規模轉移的一個例子是真核的bdelloid rotifer ,它們從細菌,真菌和植物中接收了一系列基因。病毒還可以在生物體之間攜帶DNA,即使在生物領域之間也可以轉移基因。
在獲得葉綠體和線粒體期間,真核細胞和細菌的祖先之間也發生了大規模基因轉移。真核生物本身可能起源於細菌和古細菌之間的水平基因轉移。
表觀遺傳學
某些可遺傳的變化無法通過對DNA中核苷酸序列的變化來解釋。這些現像被歸類為表觀遺傳遺傳系統。 DNA甲基化標記染色質,自我維持的代謝環,通過RNA干擾沉默的基因沉默和蛋白質的三維構象(例如prions )是在生物水平上發現表觀遺傳遺傳系統的區域。發育生物學家認為,遺傳網絡中的複雜相互作用和細胞之間的通信可能導致可遺傳的變化,這可能會構成發展可塑性和流動性方面的某些力學。遺傳力也可能在更大的尺度上發生。例如,通過生物構建過程的生態遺傳是由生物在其環境中的常規活動和重複的活動來定義的。這產生了效果的遺產,可以修改並反饋到後代的選擇方案中。不受基因直接控制的進化中遺傳力的其他例子包括文化特徵和共生髮生的遺傳。
進化力

從新達威人的角度來看,當等位基因群中的等位基因頻率變化時,例如,在繁殖生物體中的頻率發生變化,例如,在蛾類種群中,黑色的等位基因變得越來越普遍。可能導致等位基因頻率變化的機制包括自然選擇,遺傳漂移和突變偏置。
自然選擇
自然選擇的進化是在人口的後代中增強生存和繁殖的特徵變得更加普遍的過程。它體現了三個原則:
- 在形態,生理和行為(表型變化)方面,生物體種群中存在變異。
- 不同的特徵賦予不同的生存和繁殖率(差異適應性)。
- 這些特徵可以世代相傳(健身的遺傳力)。
產生的後代比可以生存的更多,這些條件會產生生存和繁殖的生物之間的競爭。因此,比那些沒有賦予優勢的特徵的生物具有特徵使他們比競爭對手具有優勢的生物更有可能將自己的特徵傳遞給下一代。這種電視是自然選擇過程創造並保留似乎適合其執行功能角色的特徵的質量。選擇的後果包括非隨機交配和遺傳搭便車。
自然選擇的核心概念是生物體的進化適應性。健身是通過生物生存和繁殖能力來衡量的,這決定了其對下一代的遺傳貢獻的大小。但是,健身與後代的總數不同:相反,健身是由隨後攜帶生物體基因的後代的比例來指示的。例如,如果生物體能夠良好生存並迅速繁殖,但是其後代太小且弱而無法生存,那麼這種生物將對子孫後代做出很少的遺傳貢獻,因此適合度較低。
如果等位基因比該基因的其他等位基因更大,那麼每個一代等位基因在人群中變得更加普遍。據說這些特徵是“選擇的”。可以提高適應性的性狀的例子是增強的生存和增加的繁殖力。相反,由於對這個等位基因的有益或有害等位基因較低或有害等位基因而造成的較低健身可能會變得更稀少 - 它們被“反對”。
重要的是,等位基因的適應性不是固定特徵。如果環境發生變化,以前中性或有害的特徵可能會變得有益,以前有益的特徵會有害。但是,即使選擇方向確實以這種方式逆轉,過去丟失的特徵也可能不會以相同的形式重新發展。但是,只要沒有從基因組中消除它們,並且只有在數百代人的基因組中被抑制,就可以重新激活這些基因,這可能會導致特徵的重新出現,被認為像牙齒一樣丟失的特徵像刺耳的牙齒一樣喪失。在雞,無翅棍昆蟲中的翅膀,人類的尾巴和其他乳頭等。這樣的“倒退”被稱為atavisms 。

人群中的自然選擇可以在高度等範圍內變化的特徵,可以歸類為三種不同類型。第一個是定向選擇,這是特徵隨時間變化的平均值的轉變,例如,生物會逐漸變高。其次,顛覆性選擇是針對極端特徵值的選擇,並且通常導致兩個不同的值變得最常見,而選擇平均值。這是當短或高的生物具有優勢,而不是中等高度的生物時。最後,在穩定選擇時,兩端都有針對極端性狀值的選擇,這會導致平均值周圍的差異降低,而多樣性則降低。例如,這將導致生物最終具有相似的高度。
自然選擇通常使自然成為個人和個體特徵或多或少生存的措施。從這個意義上講,“自然”是指生態系統,即生物系統在當地環境中與生物(物理和生物學)相互作用的系統。生態學的創始人尤金·奧杜姆( Eugene Odum系統中的材料週期(即,生存和非生存部分之間的材料交換)。...“生態系統中的每個人群都佔據了與系統其他部位不同的不同位置或位置,具有不同的關係。這些關係涉及生物體的生活史,其在食物鏈中的地位及其地理範圍。對自然的這種廣泛的理解使科學家能夠描述特定的力量,這些力量共同構成了自然選擇。
自然選擇可以在不同水平的組織中起作用,例如基因,細胞,個別生物,生物體和物種組。選擇可以同時在多個級別上起作用。選擇在單個生物水平以下的選擇的一個例子是稱為轉座子的基因,可以復制並分佈在整個基因組中。以上高於小組選擇的水平的選擇可以允許合作的演變。
遺傳漂移

遺傳漂移是從一代到下一代中人口中等位基因頻率的隨機波動。當缺乏選擇性或相對較弱時,在連續一代中,等位基因頻率同樣可能向上或向下漂移,因為等位基因會遇到採樣誤差。當等位基因最終被固定時,這種漂移停止了,要么是從人口中消失或完全替換其他等位基因。因此,遺傳漂移可能僅由於機會而從人群中消除一些等位基因。即使沒有選擇性力,遺傳漂移也會引起兩個獨立的種群,這些種群以相同的遺傳結構開頭,將不同等位基因組的不同種群分開。
根據分子進化的中性理論,大多數進化變化是通過遺傳漂移固定中性突變的結果。在此模型中,人群中的大多數遺傳變化是恆定突變壓力和遺傳漂移的結果。這種中性理論的這種形式已經進行了爭論,因為它似乎不符合自然界中看到的某些遺傳變異。該模型的一個更好支持的版本是幾乎中立的理論,根據該理論,在較小人群中,在較小人群中有效中性的突變在大人群中不一定是中性的。其他理論表明,遺傳漂移在進化中被其他隨機力量縮短,例如遺傳搭式班班,也稱為遺傳草稿。另一個概念是建設性的中性進化(CNE),它解釋說,由於過多的容量,預抑制和棘輪的原則,複雜的系統可以通過中性過渡出現並擴散到人群中,並且它已應用於從起源於起源於與微生物群落複雜相互依賴性的剪接。
被遺傳漂移固定的中性等位基因所需的時間取決於人口規模。較小的人群中的固定更快。人口中的個體數量並不重要,而是一種稱為有效人口規模的措施。有效人群通常小於總人口,因為它考慮了諸如近親繁殖水平和人口最小的生命週期的階段。在同一人群中,每個基因的有效人口規模可能並不相同。
通常很難衡量選擇和中性過程(包括漂移)的相對重要性。適應性和非自適應力在推動進化變化中的比較重要性是當前研究的一個領域。
突變偏見
突變偏差通常被認為是兩種不同類型的突變,例如,過渡轉換偏置,GC-AT偏置,缺失插入偏置的預期速率差異。這與發展偏見的觀念有關。 Haldane和Fisher認為,由於突變很容易通過選擇而克服,因此突變趨勢將是無效的,除非在中性進化或非常高的突變率的條件下。長期以來,這種相反的壓力論點被用來駁斥進化中內部趨勢的可能性,直到分子時代促使人們對中性進化產生了興趣。
Noboru Sueoka和Ernst Freese提出,突變中的系統偏見可能導致物種之間基因組GC組成的系統差異。 1967年,鑑定出GC偏向大腸桿菌突變菌株的鑑定,以及中性理論的提議,確定了分子模式的突變解釋的合理性,這些解釋現在在分子進化文獻中很常見。
例如,在密碼子使用模型中經常調用突變偏差。這樣的模型還包括選擇的效果,遵循突變選擇 - 拖船模型,該模型允許基於翻譯的影響來實現突變偏見和差異選擇。突變偏差的假設在思考基因組組成的演變的發展中起著重要作用,包括等距。不同分類單元中不同的插入與缺失偏見可以導致不同基因組大小的演變。關於基因組大小的林奇的假設依賴於突變偏見,以增加或減少基因組大小。
然而,當發現(1)GC偏見的基因轉化率在二倍體生物(例如哺乳動物)和(2)細菌基因組中經常偏見時,對組成演變的突變假設卻降低了範圍的降低。突變。
當代思考突變偏見的作用反映了與霍爾丹和費舍爾的理論不同的理論。最新的工作表明,原始的“壓力”理論假設進化是基於站立的變化:當進化取決於突變事件時,引入新等位基因,突變和發育偏見的引入變化(到達偏見)可能對進化構成偏見。不需要中性進化或高突變率。幾項研究報告說,與適應有關的突變反映了常見的突變偏見,而其他突變則對此解釋產生了異議。
遺傳搭便車
重組使同一條DNA上的等位基因分離。但是,重組率很低(每一代染色體大約兩個事件)。結果,在染色體上緊密的基因可能並不總是彼此脫落,而結合在一起的基因往往會遺傳在一起,這是一種稱為鏈接的現象。通過發現兩個等位基因在單個染色體上與期望相比,這被稱為其連鎖不平衡的頻率來衡量這種趨勢。一組通常在組中繼承的等位基因稱為單倍型。當特定單倍型中的一個等位基因非常有益時,這可能很重要:自然選擇可以推動選擇性掃描,這也會導致單倍型中的其他等位基因在人群中變得越來越普遍;這種效果稱為遺傳搭便車或遺傳草稿。遺傳草稿是由於某些中性基因與正在選擇的其他基因相關的事實引起的,可以部分通過適當的有效種群規模來部分捕獲。
性選擇

自然選擇的一種特殊情況是性選擇,這是通過提高生物體對潛在伴侶的吸引力來增加交配成功的任何特徵的選擇。通過性選擇進化的特徵在幾種動物物種的雄性中尤為突出。儘管受到性青睞,但諸如繁瑣的鹿角,交配呼叫,大體型和鮮豔的色彩之類的特徵通常會吸引捕食,從而損害了個體男性的生存。這種生存劣勢是通過表現出這些難以欺騙的性特徵的男性的更高生殖成功來平衡的。
自然結果
進化會影響生物體形式和行為的各個方面。最突出的是自然選擇結果的特定行為和身體適應。這些適應通過協助尋找食物,避免捕食者或吸引伴侶來增加健身。有機體還可以通過相互協作,通常是通過協助其親戚或從事互惠互利共生的方法來應對選擇。從長遠來看,進化通過將祖先的生物種群分裂成新的群體,從而產生新物種,這些群體無法或不會雜交。這些進化的結果是根據時間尺度作為宏觀進化與微發展的。宏觀進化是指在物種水平或高於物種水平的進化中,特別是物種形成和滅絕。而微發展是指物種或人群內的進化變化較小,特別是等位基因頻率和適應性變化。宏觀進化的小型進化的結果。因此,微觀進化和宏觀進化之間的區別不是基本的 - 差異僅僅是涉及的時間。但是,在宏觀進化中,整個物種的特徵可能很重要。例如,個體之間的大量差異使一個物種可以快速適應新的棲息地,減少了它滅絕的機會,而廣泛的地理範圍會增加物種形成的機會,從而使部分人口的一部分更有可能會增加變得孤立。從這個意義上講,微觀進化和宏觀進化可能涉及在不同級別的選擇,該級別具有作用於基因和生物的微觀進化,與宏觀進化過程相比,例如作用於整個物種的物種選擇並影響其物種形成和滅絕的速度。
一個普遍的誤解是,進化具有目標,長期計劃或天生的“進步”傾向,如正性和進化論諸如信念所表達的那樣;但是,實際上,進化沒有長期目標,不一定會產生更大的複雜性。儘管複雜的物種已經發展,但它們的出現是生物總數增加的副作用,而簡單的生命形式在生物圈中仍然更為普遍。例如,絕大多數物種是微觀原核生物,儘管大小很小,但它們構成了世界上一半的生物量,並且構成了地球絕大多數生物多樣性。因此,簡單的生物在整個歷史上一直是地球上生命的主要形式,並且一直是迄今為止生命的主要形式,複雜的生活只會更加多樣化,因為它更加明顯。確實,微生物的進化對於進化研究尤為重要,因為它們的快速繁殖允許研究實驗進化以及實時觀察進化和適應。
適應

適應是使生物更適合其棲息地的過程。同樣,改編一詞可能是指對於生物的生存至關重要的特徵。例如,馬的牙齒適應草的磨碎。通過將術語適應用於產品的進化過程和自適應性狀(身體部分或功能),可以區分單詞的兩種感覺。適應是由自然選擇產生的。以下定義是由於Theodosius Dobzhansky造成的:
- 適應性是進化過程,生物體變得更好地生活在其棲息地或棲息地中。
- 適應性是適應的狀態:生物體能夠在給定的一組棲息地中生活和繁殖的程度。
- 自適應特徵是生物體發育模式的一個方面,它可以使或增強該生物的生存和繁殖的可能性。
適應可能會導致新功能的增益或祖先特徵的喪失。一個顯示兩種類型變化的例子是細菌適應抗生素選擇,遺傳變化通過修飾藥物的靶標或增加將藥物從細胞中泵出的轉運蛋白的活性來引起抗生素耐藥性。其他引人注目的例子是細菌大腸桿菌在長期實驗室實驗中使用檸檬酸作為養分的能力,黃桿菌進化出一種新型酶,使這些細菌可以在尼爾龍製造的副產品和土壤細菌的副產品上生長鞘磷脂進化出一種全新的代謝途徑,該途徑降解了合成農藥五氯苯酚。一個有趣但仍然有爭議的想法是,某些適應可能會提高生物體產生遺傳多樣性並通過自然選擇(增加生物體的可發展性)適應的能力。
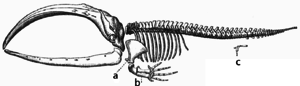
適應是通過逐漸修改現有結構進行的。因此,具有相似內部組織的結構在相關生物中可能具有不同的功能。這是單個祖先結構以不同方式調整功能的結果。例如,由於普通哺乳動物祖先的所有這些結構的下降,蝙蝠翅膀內的骨頭與小鼠腳和靈長類動手的骨頭非常相似。但是,由於所有生物在某種程度上都是相關的,即使似乎幾乎沒有或沒有結構性相似性的器官,例如節肢動物,魷魚和脊椎動物的眼睛,或者是節肢動物和脊椎動物的四肢和翅膀,也可以取決於一組常見的集合控制其組裝和功能的同源基因;這稱為“深層同源” 。
在進化過程中,某些結構可能會失去其原始功能並成為殘留結構。此類結構在當前物種中可能幾乎沒有功能,但在祖先物種或其他密切相關的物種中具有明顯的功能。例子包括偽基因,眼睛中的非功能性遺體,無飛鳥的翅膀,鯨魚和蛇中的髖骨存在以及通過無性繁殖繁殖的生物體中的性特徵。人類殘留結構的例子包括智慧牙齒, Coccyx , Vermiform附錄以及其他行為遺跡,例如鵝腫塊和原始反射。
但是,許多似乎是簡單適應的特徵實際上是顯著的:最初適用於一個函數的結構,但巧合的是,在此過程中對某些其他功能有些有用。一個例子是非洲蜥蜴霍斯皮斯·蓋特里(Lizard Holaspis Guentheri) ,它開發了一個非常平坦的頭部,可以隱藏在縫隙中,可以看出靠近的親戚。但是,在該物種中,頭部變得如此扁平,以至於它有助於從樹上滑行,這是一種消除。在細胞中,通過募集幾種以前具有不同功能的預先存在的蛋白質的募集,分子機器(例如細菌鞭毛和蛋白質分類機械)演變出來。另一個例子是從糖酵解和異種代謝中募集酶,用作有機體眼鏡中稱為結晶蛋白的結構蛋白。
當前在進化發育生物學中進行調查的領域是適應和應對的發展基礎。這項研究探討了胚胎髮育的起源和演變以及發展和發展過程的修改如何產生新的特徵。這些研究表明,進化可以改變發育以產生新的結構,例如胚胎骨結構在其他動物的頜骨中發展成頜骨,而不是在哺乳動物中形成中耳的一部分。由於發育基因的變化,例如在進化中丟失的結構也可能會重新出現,例如雞的突變導致胚胎長出類似於鱷魚的牙齒。現在越來越清楚的是,生物體形式的大多數改變是由於一小部分保守基因的變化所致。
協同進化

生物之間的相互作用可以產生衝突和合作。當相互作用是在病原體和宿主或捕食者及其獵物等物種對之間的相互作用時,這些物種可以產生匹配的適應集。在這裡,一個物種的演變導致第二種適應。然後,第二種物種的這些變化又導致了第一個物種的新適應。這種選擇和響應的周期稱為協同進化。一個例子是在粗糙的NEWT中產生四毒素,以及其捕食者中的四毒素耐藥性的進化,即公共吊蛇蛇。在這對捕食者捕食對中,進化的武器競賽在NEWT中產生了高水平的毒素,並且在蛇中相應的高水平毒素抗性。
合作
並非所有物種之間的共同進化相互作用都涉及沖突。許多互惠相互作用的病例已經發展。例如,植物與菌根真菌之間存在極端的合作,它們在其根部生長,並幫助植物從土壤中吸收營養。這是一種相互關係,因為植物為真菌提供了來自光合作用的糖。在這裡,真菌實際上生長在植物細胞內,使它們能夠與宿主交換營養,同時發送抑制植物免疫系統的信號。
同一物種的生物之間的聯盟也在發展。一個極端的情況是在社會昆蟲(例如蜜蜂,白蟻和螞蟻)中發現的歐洲社會性,在該昆蟲中,無菌昆蟲在一個能夠繁殖的菌落中少量的生物體餵養和守護了少量的生物。在較小的規模上,構成動物身體的體細胞限制了它們的繁殖,因此它們可以保持穩定的生物體,然後支持少數動物的生殖細胞產生後代。在這裡,體細胞會響應特定信號,這些信號指示他們是否成長,保持原狀或死亡。如果細胞忽略這些信號並不合時宜地倍增,則它們的不受控制的生長會導致癌症。
物種內部的這種合作可能是通過選擇親屬的過程而發展的,這是一個生物的作用來幫助提高親戚的後代。之所以選擇此活動,是因為如果幫助個體包含促進幫助活動的等位基因,則其親屬也可能包含這些等位基因,因此這些等位基因將被傳遞。可能促進合作的其他過程包括小組選擇,合作為一組生物帶來好處。
物種形成

物種是一個物種分解為兩個或多個後代物種的過程。
有多種方法可以定義“物種”的概念。定義的選擇取決於相關物種的特殊性。例如,某些物種概念更容易地應用於性生殖生物,而另一些物種概念則更好地傾向於無性無性生物。儘管各種物種概念的多樣性,但這些各種概念都可以置於三種廣泛的哲學方法之一中:雜交,生態和系統發育。生物物種概念(BSC)是雜交方法的經典例子。 BSC在1942年由進化生物學家恩斯特·梅爾(Ernst Mayr)定義,“物種是實際或潛在雜交的天然種群的群體,它們與其他此類群體的生殖分離。”儘管具有廣泛和長期的使用,但像其他物種概念一樣,BSC並非沒有爭議,因為原核生物之間的遺傳重組不是生殖的內在方面。這稱為物種問題。一些研究人員試圖對物種進行統一的統一定義,而另一些研究人員則採用了多元化方法,並認為可能有不同的方法可以從邏輯上解釋物種的定義。
需要兩個不同性種群之間繁殖的障礙才能成為新物種。基因流也可以通過將新的遺傳變體傳播到其他種群來減慢這一過程。根據自最近的共同祖先以來兩個物種的分歧,他們仍然有可能產生後代,就像馬和驢交配一樣產生mu子。這種雜種通常是不育的。在這種情況下,與密切相關的物種可能定期雜交,但將選擇雜種,並且該物種將保持不同。但是,偶爾會形成可行的雜種,這些新物種可以在其父物種之間具有中間的特性,或者俱有全新的表型。雜交在生產新動物中的重要性尚不清楚,儘管在許多類型的動物中都發現了病例,而灰樹青蛙是一個特別好的例子。
在受控實驗室條件和自然界中,已經觀察到了多次物種形成。在性繁殖生物中,物種形成是由生殖隔離而產生的,其次是譜系差異。物種形成的主要地理模式有四種。動物中最常見的是同種異體形成,它發生在最初在地理上孤立的人群中,例如棲息地破碎或遷移。在這些條件下的選擇會導致生物的外觀和行為發生非常快速的變化。隨著選擇和漂移獨立於與其其他物種隔離的種群中,分離最終可能會產生無法雜交的生物。
第二種形態的模式是圍存在形成,這是在新環境中孤立的小種群體時發生的。這與同種異體形成不同,因為孤立的人群在數值上比父母種群小得多。在這裡,創始人的效應在增加近交之後會導致近交之後的快速形態,從而導致遺傳變化迅速。
第三種模式是伴侶形成。這類似於圍牙形成,因為一個人群進入了一個新的棲息地,但不同之處在於,這兩個人群之間沒有物理分離。取而代之的是,物種形成是由減少兩個種群之間基因流動的機制的演變而產生的。通常,當父母物種棲息地內的環境發生巨大變化時,這會發生這種情況。一個例子是草果鳥th蟲,它可以響應於礦山的局部金屬污染而經歷屬性物種。在這裡,植物進化,對土壤中高水平的金屬具有抗性。與金屬敏感的父母種群相互雜交的選擇會導致抗金屬植物的開花時間逐漸變化,這最終產生了完全的生殖隔離。針對兩個種群之間的雜種的選擇可能會導致增強,這是促進一個物種內交配的性狀的演變以及特徵位移,這是兩個物種在外觀上變得更加獨特的時候。

最後,在同胞形成物種中,沒有地理隔離或棲息地變化而發散。這種形式很少見,因為即使少量基因流也可能消除人群部分之間的遺傳差異。通常,動物的同胞形成需要遺傳差異和非隨機交配的演變,以允許生殖分離進化。
一種類型的同胞形成涉及兩個相關物種的雜交,以產生新的雜種物種。這在動物中並不常見,因為動物雜種通常是無菌的。這是因為在減數分裂期間,來自每個父母的同源染色體來自不同的物種,無法成功配對。但是,在植物中更常見,因為植物通常會增加染色體的數量,形成多倍體。這使每個父母物種的染色體在減數分裂過程中形成匹配對,因為每個父母的染色體已經由一對錶示。這樣的物種事件的一個例子是,植物種類擬南芥和擬南芥的植物雜交雜種以賦予新物種擬南芥suecica 。這發生在大約20,000年前,並且在實驗室中重複了物種過程,該過程允許研究此過程中涉及的遺傳機制。實際上,在一個物種中加倍的染色體可能是生殖隔離的常見原因,因為在育種有生物體時,將無與倫比的一半染色體是無與倫比的。
物種事件在標點平衡的理論中很重要,該理論解釋了散佈在相對較長的停滯時期的變化“爆發”的化石記錄中的模式,那裡的物種保持相對不變。在這一理論中,物種形成和快速進化是聯繫的,自然選擇和遺傳漂移最強烈地影響了新型棲息地或小種群中種種物種的生物。結果,化石記錄中的停滯週期對應於父母種群,並且在小種群或地理限制的棲息地中發現了經歷物種形成和快速進化的生物,因此很少被保留為化石。
滅絕

滅絕是整個物種的消失。滅絕並不是異常事件,因為物種經常通過物種形成出現並通過滅絕消失。現在幾乎所有生活在地球上的動物和植物物種都滅絕了,滅絕似乎是所有物種的最終命運。儘管偶爾發生的大規模滅絕事件中的滅絕率速率,但這些滅絕的發生在整個生命的歷史中一直不斷發生。白堊紀 - 銷去滅絕事件在此期間是最著名的,但較早的二疊紀 - 三疊紀滅絕事件更加嚴重,大約96%的海洋物種被滅絕。全新世滅絕事件是過去幾千年來人類在全球範圍內擴張的持續巨大滅絕。當前的滅絕率是背景速率的100–1000倍,到21世紀中葉,當前物種的30%可能已滅絕。現在,人類活動是持續滅絕事件的主要原因;將來,全球變暖可能會進一步加速它。儘管估計在地球上居住的所有物種中有超過99%的滅絕,但估計約有1萬億種在地球上,目前僅描述了1%的1%。
滅絕在進化中的作用不是很好地理解,並且可能取決於考慮哪種類型的滅絕類型。構成大多數滅絕的連續“低級”滅絕事件的原因可能是物種之間競爭有限資源(競爭性排除原則)的結果。如果一種物種可以兼容另一個物種,則可以產生物種的選擇,而更矮的物種存活,而其他物種則被滅絕。間歇性的巨大滅絕也很重要,但是它們並沒有充當選擇性的力量,而是以非特異性的方式大大降低了多樣性,並促進了倖存者快速發展和物種形成的爆發。
申請
進化生物學中使用的概念和模型(例如自然選擇)具有許多應用。
人工選擇是有機體中有意選擇特徵。這已經用於植物和動物的馴化已有數千年了。最近,這種選擇已成為基因工程的重要組成部分,可選標記(例如使用抗生素抗性基因來操縱DNA)。具有寶貴特性的蛋白質通過在稱為有向進化的過程中反復進行突變和選擇(例如改良的酶和新抗體)而發展。
了解生物體進化過程中發生的變化可以揭示構造身體部分所需的基因,這些基因可能涉及人類遺傳疾病。例如,墨西哥四人是一種白化洞魚,在進化過程中失去了視力。將這種盲魚的不同種群繁殖在一起產生了一些用功能性的眼睛的後代,因為在不同洞穴中進化的孤立種群發生了不同的突變。這有助於確定視力和色素沉著所需的基因。
進化論在醫學中有許多應用。許多人類疾病不是靜態現象,而是能夠進化的現象。病毒,細菌,真菌和癌症能夠抗宿主免疫防禦和藥物。這些同樣的問題在農藥和除草劑耐藥性中發生。我們可能面臨大多數可用抗生素的有效壽命的終結,並預測病原體的演變和發展性,並製定策略以減慢或規避,這需要更深入地了解在分子水平上推動進化的複雜力的知識。
在計算機科學中,使用進化算法和人工生命的進化模擬始於1960年代,並通過人工選擇的模擬進行了擴展。由於Ingo Rechenberg在1960年代的工作,人工進化成為一種廣泛認可的優化方法。他使用進化策略來解決複雜的工程問題。遺傳算法尤其是通過約翰·亨利·霍蘭德(John Henry Holland)的撰寫而受歡迎的。實際應用還包括計算機程序的自動演變。現在,進化算法用於比人類設計師生產的軟件更有效地解決多維問題,並優化系統的設計。
生命的進化史
生命的起源
地球約為45.4億年。地球生命最早的無可爭議的證據的歷史可以追溯到至少35億年前,即在較早的熔融Hadean Eon之後的地質地殼開始鞏固之後的Eoarchean時代。在西澳大利亞州有34.8億年曆史的砂岩中發現了微生物墊化石。在西部格陵蘭島發現的37億年曆史的生物物質的其他早期物理證據是石墨,以及在西澳大利亞州41億年曆史的岩石中發現的“生物生物遺體”。斯蒂芬·布萊爾·海奇斯(Stephen Blair Hedges)在評論澳大利亞的發現時寫道:“如果生命在地球上相對較快,那麼在宇宙中可能很普遍。” 2016年7月,科學家報導說,從地球上所有生物體的所有生物體中,從最後一個普遍的共同祖先(LUCA)中識別出一組355個基因。
據估計,所有物種中有超過50億種的物種中有99%以上的物種已滅絕。關於地球當前物種數量的估計量從1000萬到1400萬,其中約有190萬個被命名為160萬,迄今為止,中央數據庫中記錄了160萬,至少尚未描述80%。
人們認為高能化的化學作用大約在40億年前產生了一個自我複制的分子,而十億年後,所有生命的最後一個共同的祖先就存在。當前的科學共識是,構成生命的複雜生物化學來自更簡單的化學反應。生命的開始可能包括自我複制分子,例如RNA和簡單細胞的組裝。
常見下降
地球上的所有生物都來自共同的祖先或祖先基因庫。當前物種是進化過程中的一個階段,其多樣性是一系列形成和滅絕事件的產物。有機體的常見下降首先是從有關生物體的四個簡單事實中得出的:首先,它們具有無法通過局部適應來解釋的地理分佈。其次,生命的多樣性不是一組完全獨特的生物,而是具有形態學相似性的生物。第三,沒有明確目的的殘留性狀類似於功能性祖先特徵。第四,可以使用這些相似性將生物分類為類似於家譜的嵌套組的層次結構。
由於水平基因的轉移,這種“生命之樹”可能比簡單的分支樹更為複雜,因為某些基因已在遠距離相關的物種之間獨立傳播。為了解決這個問題,有些作者更喜歡將“生命的珊瑚”用作隱喻或數學模型來說明生命的演變。這種觀點可以追溯到達爾文簡短提到的一個想法,但後來放棄了。
過去的物種也留下了其進化史的記錄。化石以及當今生物的比較解剖結構構成形態學或解剖學記錄。通過比較現代和滅絕物種的解剖學,古生物學家可以推斷這些物種的譜系。但是,對於具有硬體部位的生物(例如殼,骨頭或牙齒),這種方法最成功。此外,由於細菌和古細菌等原核生物具有有限的共同形態,因此它們的化石無法提供有關其祖先的信息。
最近,共同下降的證據來自生物之間生化相似性的研究。例如,所有活細胞使用相同的基本核苷酸和氨基酸。分子遺傳學的發展揭示了生物體基因組中留下的進化記錄:何時在突變產生的分子時鐘差異時約會。例如,這些DNA序列比較表明,人類和黑猩猩共享98%的基因組,並分析它們不同的少數區域,這有助於闡明這些物種的共同祖先的存在。
生命的進化

原核生物大約在3-4億年前居住在地球上。在接下來的幾十億年中,這些生物體中沒有明顯的形態或細胞組織發生明顯變化。真核細胞在1.6至27億年前出現。細胞結構的下一個重大變化是當細菌被真核細胞吞沒的,這是一個稱為內共生的合作關聯。然後,吞噬的細菌和宿主細胞進行了共同進化,細菌進化為線粒體或氫化體。藍細菌樣生物的另一種吞噬導致藻類和植物中的葉綠體形成。
生命的歷史是單細胞真核生物,原核生物和古細菌的歷史,直到大約6.1億年前,當時多細胞生物在埃迪亞卡蘭時期開始出現在海洋中。多細胞的演變發生在多個獨立事件中,例如海綿,棕色藻類,藍細菌,粘液黴菌和粘菌病的生物。 2016年1月,科學家報告說,大約8億年前,一種稱為GK-PID的單個分子的遺傳變化可能使生物體從單個細胞生物體變成了許多細胞之一。
在這些第一個多細胞生物出現後不久,在稱為坎布里亞爆炸的事件中,大約1000萬年出現了顯著的生物多樣性。在這裡,大多數現代動物出現在化石記錄中,以及隨後滅絕的獨特譜系。已經提出了寒武紀爆炸的各種觸發因素,包括從光合作用中氧氣在大氣中的積累。
大約5億年前,植物和真菌殖民了這片土地,隨後是節肢動物和其他動物。昆蟲特別成功,即使在今天,大多數動物物種也構成了大多數動物。兩棲動物首次出現在大約364億年前,其次是早期的羊膜和鳥類,大約1.55億年前(均來自“爬行動物”類似譜系),哺乳動物大約1.29億年前,大約1000萬年前,現代人類和250,000年的現代人類前。然而,儘管這些大動物的發展,但較小的生物體類似於此過程中早期進化的類型,仍然非常成功並佔據地球的主導地位,大多數生物量和物種都是原核生物。
進化思想的歷史




古典古代
一種有機體可以從另一種類型中降下來的提議可以追溯到一些前op前的希臘哲學家,例如Anaximander和Empedocles 。這樣的建議倖存到羅馬時代。詩人兼哲學家盧克雷烏斯(Lucretius)在他的傑作《 de rerum natura》中跟隨Empedocles(點燃。 “關於事物的本質”)。
中世紀
與這些唯物主義的觀點相反,亞里士多德主義將所有自然事物視為固定自然可能性的實現,稱為形式。這成為中世紀對自然的目的論理解的一部分,其中所有事物都具有神聖宇宙秩序的意圖作用。這個想法的變化成為了中世紀的標準理解,並被融入了基督教的學習中,但是亞里士多德並不要求真正的生物類型的生物總是與確切的形而上學形式一對一地對應,並專門舉例說明了新型生活類型的例子事情可能是。
許多阿拉伯穆斯林學者寫了關於進化論的文章,最著名的是伊本·哈爾登(Ibn Khaldun ),他在公元1377年寫了穆卡迪瑪(Muqaddimah)的書,他斷言,人類是從“猴子世界”中發展而來的,在一個過程中,“物種變得越來越多,” ”。
darwinian前
17世紀的“新科學”拒絕了亞里士多德的方法。它試圖用物理定律來解釋自然現象,這些現像對於所有可見的事物都是相同的,並且不需要任何固定的自然類別或神聖的宇宙秩序。但是,這種新方法紮根於生物科學:固定自然類型概念的最後堡壘。約翰·雷(John Ray)應用於固定自然類型的“物種”和動物類型的固定類型“物種”,但他將每種類型的生物識別為一種物種,並提出每個物種可以由持續的特徵定義自己一代一代。卡爾·林納斯(Carl Linnaeus)在1735年引入的生物學分類明確識別了物種關係的等級性質,但仍將物種視為根據神聖計劃固定的。
這段時間的其他博物學家推測物種隨時間的進化變化,根據自然法則。 1751年,皮埃爾·路易斯·莫珀圖斯(Pierre Louis Maupertuis)寫道,在繁殖過程中發生了自然修飾,並在許多世代積累了生產新物種。喬治 - 路易斯·萊克萊克(Georges-Louis Leclerc),孔德·德布朗(Comte de Buffon )建議物種可以退化為不同的生物體,伊拉斯mus達爾文(Erasmus darwin)提出,所有溫血動物都可以從單一的微生物(或“絲狀”)中降下。第一個成熟的進化方案是讓·巴蒂斯特·拉馬克(Jean-Baptiste Lamarck)的“ trans變” 1809年理論,該理論設想自發產生不斷產生簡單的生活形式,在平行血統中具有固有的漸進式趨勢,並假定在局部趨勢上,並假定這些血統通過繼承父母使用或廢棄的變化來適應環境。 (後一個過程後來被稱為拉馬克主義。)這些思想被既定的博物學家譴責,因為猜測缺乏經驗支持。喬治(Georges)尤其是庫維耶(Georges)堅持認為物種是無關的和固定的,它們的相似之處反映了神聖的設計滿足了功能需求。同時,威廉·佩利(William Paley)發展了雷的仁慈設計思想,它是對神的存在和屬性的自然神學或證據(1802)的發展,該神學和屬性提出了複雜的改編作為神聖設計的證據,並受到查爾斯·達爾文(Charles Darwin)的欽佩。
達爾文革命
恆定類型學類別或生物學類型的概念的關鍵突破是通過自然選擇的進化論,這是由查爾斯·達爾文(Charles Darwin)和阿爾弗雷德·華萊士( Alfred Wallace)以可變人群的形式提出的。達爾文使用了“帶有修飾的下降”而不是“進化”。達爾文(Darwin)受到托馬斯·羅伯特·馬爾薩斯(Thomas Robert Malthus )的人口原則(1798)的一篇文章的影響,人口增長將導致“生存鬥爭”,其中有利的變化隨著其他人的滅亡而佔了上風。在每一代人中,由於資源有限,許多後代無法生存到繁殖時代。這可以解釋從共同血統到自然法律的多樣性,以相同的方式對所有類型的生物來說。達爾文(Darwin)從1838年開始就發展了他的“自然選擇理論”理論,並在阿爾弗雷德·羅素·華萊士(Alfred Russel Wallace)向他發送了幾乎相同理論的版本時,寫了他的“ 大書”。倫敦林尼亞學會。 1859年底,達爾文(Darwin)出版了他的“摘要”,詳細解釋了自然選擇,並以一種越來越多地接受了達爾文進化的概念,以犧牲替代理論為代價。托馬斯·亨利·赫x黎(Thomas Henry Huxley)使用古生物學和比較解剖學將達爾文的思想應用於人類,以提供有力的證據,表明人類和猿類共有一個共同的血統。有些人對此感到不安,因為它暗示人類在宇宙中沒有特殊的位置。
腫瘤和遺傳
生殖遺傳力和新特徵的起源的機制仍然是一個謎。為此,達爾文發展了他的pangenesis臨時理論。 1865年,格雷戈爾·門德爾(Gregor Mendel)報告說,特徵是通過獨立的元素分類和隔離(後來稱為基因)以可預測的方式遺傳的。孟德爾的繼承定律最終取代了達爾文的大部分pangenesis理論。八月韋斯曼(August Weismann)在產生配子(例如精子和卵細胞)的生殖細胞和人體的體細胞之間做出了重要的區別,表明遺傳僅通過了細菌。 Hugo de Vries將Darwin的Pangenesis理論與Weismann的細菌/SOMA細胞區別聯繫在一起,並提出Darwin的Pangenes集中在細胞核中,當表達時,它們可以移入細胞質以改變細胞的結構。德·弗里斯(De Vries)也是使門德爾(Mendel)作品眾所周知的研究人員之一,他認為孟德爾(Mendelian)的特徵與沿著種系的可遺傳變異轉移相對應。為了解釋新變體是如何產生的,de vries開發了一種突變理論,導致那些接受達爾文進化論的人與與de vries結盟的生物有限公司之間的暫時裂痕。在1930年代,人口遺傳學領域的開拓者,例如羅納德·費舍爾(Ronald Fisher) ,塞瓦爾·賴特(Sewall Wright )和賈布斯·哈爾丹(JBS Haldane),將進化的基礎設置為強大的統計哲學。因此,調和了達爾文理論,基因突變和孟德爾的繼承之間的錯誤矛盾。
“現代綜合”
在1920年代和1930年代,現代合成將基於孟德爾遺傳的自然選擇和種群遺傳學聯繫在一起,包括一個統一的理論,其中包括隨機遺傳漂移,突變和基因流動。這種新版本的進化論重點是人口中等位基因頻率的變化。它通過古生物學中的化石過渡解釋了種群中種群中觀察到的模式。
進一步的合成
從那時起,根據許多發現,進一步的合成擴大了進化的解釋能力,以涵蓋從基因到種群的整個生物層次結構的生物學現象。
詹姆斯·沃森(James Watson )和弗朗西斯·克里克( Francis Crick)在1953年貢獻了詹姆斯·沃森(James Watson)和弗朗西斯·克里克(Francis Crick)的DNA結構的出版,證明了繼承的物理機制。分子生物學改善了對基因型與表型之間關係的理解。系統發育系統學也取得了進步,通過發表和使用進化樹將特徵的過渡映射到比較和可檢驗的框架中。 1973年,進化生物學家西奧多斯·多布津斯基(Theodosius Dobzhansky)寫道:“除了進化之外,生物學上沒有任何意義,因為它已經揭示了最初在自然歷史上首先脫節的事實的關係,成為描述和描述和描述和描述的知識體系。預測了許多關於這個星球生命的可觀察到的事實。
一種擴展,稱為進化發育生物學,非正式地稱為“ evo-devo”,強調了世代(進化)(進化)之間的變化如何對單個生物體內部變化模式作用(開發)。自21世紀初以來,一些生物學家主張擴展的進化綜合,這將解釋非遺傳遺傳模式的影響,例如表觀遺傳學,父母效應,生態遺傳和文化遺傳和可發展性。
社會和文化反應

在19世紀,尤其是在1859年出版《物種起源》之後,生命發展的觀念是積極的學術辯論來源,以進化的哲學,社會和宗教含義為中心。如今,現代的進化綜合已被絕大多數科學家所接受。但是,對於某些有神論者來說,進化仍然是一個有爭議的概念。
儘管各種宗教和教派通過諸如有神論的進化之類的概念使他們的信仰與進化一致,但有一些創造論者認為進化與他們宗教中的創造神話相矛盾,並提出了對進化的各種反對意見。正如對1844年自然創造自然歷史遺跡的回應所證明的那樣,進化生物學的最具爭議的方面是人類進化的含義,即人類與猿猴共同的祖先和人類人類精神和道德能力的共同祖先具有與動物中其他遺傳性狀相同類型的自然原因。在某些國家,尤其是美國,科學與宗教之間的這些緊張局勢加劇了當前的創造與進化的爭議,這是一場宗教衝突,重點是政治和公共教育。儘管諸如宇宙學和地球科學等其他科學領域也與許多宗教文本的文字解釋相抵觸,但進化生物學經歷的經歷更加多地反對宗教文字主義者。
在20世紀上半葉的大部分時間裡,美國中學生物學課程的進化教學並不常見。 1925年的範圍審判決定使該受試者在一代人的美國二級生物學教科書中變得非常罕見,但稍後逐漸重新引入,並受到1968年Epperson訴Arkansas訴Arkansas判決的法律保護。從那時起,在1970年代和1980年代的各種決定中,在中學課程中,具有競爭性的創造論的宗教信仰在法律上不允許,但它以偽科學的形式返回,為智能設計( ID),在2005年Kitzmiller訴Dover訴Dover案中再次被排除在外地區學區案例。關於達爾文思想的辯論在中國沒有引起重大爭議。
也可以看看
- 權力下放(生物學) - 物種可以恢復為原始形式的觀念